
Genetisk variation betraktas som en nivå av biologisk mångfald. Det är ett mått på hur genetiskt olika individer är inom en hel art eller en population av en art. Med Konventionen om biologisk mångfald, som är en del av FN:s miljöprogram, har Sverige liksom 195 andra länder förbundit sig att bevara den biologiska mångfalden i landet på ekosystem-, art- och gennivå.
Genetisk variation är grunden för evolution och anpassning. Om alla individer inom en art eller population är genetiskt lika kommer alla individer att vara känsliga för samma förändringar i miljön. Ett nytt virus eller ett varmare klimat skulle då slå hårt mot hela gruppen. Med en högre grad av genetisk variation ökar chansen att några individer har just de egenskaper som krävs för att anpassa sig till en förändrad livsmiljö. Det är därför en slags försäkring för artens, eller populationens, överlevnad.
Mutationer, sexuell förökning och att det sker ett genetiskt utbyte mellan populationer leder till en högre genetisk variation. Faktorer som däremot minskar den genetiska variationen är genetisk drift, flaskhals- och grundareffekter.
Faktorer som ökar genetisk variation
Mutationer är förändringar i DNA-sekvensen som uppkommer spontant i alla livsformer och kan ge upphov till helt nya egenskaper. Om mutationen ökar bärarens chanser att överleva och föröka sig kan den över tid spridas och bli vanlig i en population. Hos arter med sexuell reproduktion måste mutationen finnas i en könscell för att kunna föras vidare till nästa generation. När mutationer uppkommer i somatiska celler, ärvs de visserligen vidare till andra celler vid celldelningen, men stannar i individen. Det kallas för somatiska mutationer.

Kopieringsfel leder till mutationer
En cell delar sig ett antal gånger under sin livstid. På så sätt bildas hela tiden nya celler i kroppen som kan ersätta gamla eller skadade celler. Inför varje celldelning kopieras allt DNA så att det i alla celler finns en kopia av individens genom. Ibland uppstår det fel vid kopieringen. Oftast lyckas cellen laga det själv, men vid ungefär var tionde celldelning blir felet bestående. En mutation har då uppkommit.
Om DNA exponeras för vissa kemikalier eller viss strålning kan det skadas. På samma sätt som vid kopieringsfelen försöker cellerna att laga skadorna som uppkommit, och när det misslyckas uppstår mutationer. Det finns också en viss typ av virus, så kallade retrovirus, som fogar in sitt DNA på flera platser i värdcellens genom och därmed orsakar mutationer.
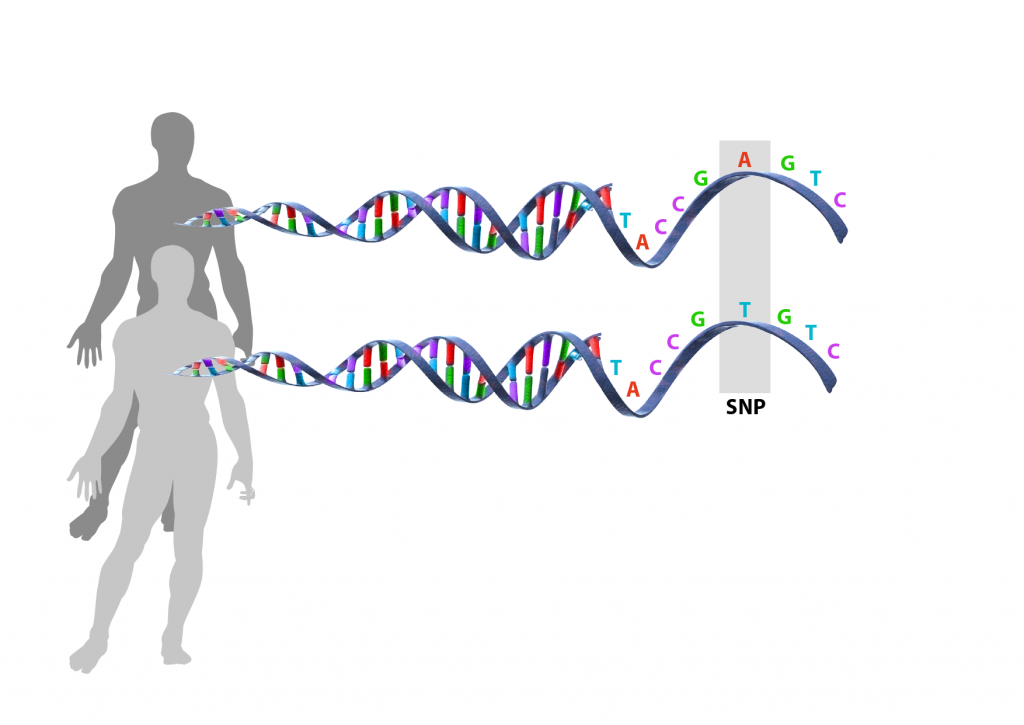
Olika sorters mutationer
En mutations effekt beror bland annat på vilken sorts mutation det är och var i genomet den uppstår. Vissa typer av mutationer ”rör om” mer än andra.
- Enbaspolymorfier är små substitutioner som till exempel kan innebära att basparet A-T förändras till C-G. I Sverige används ofta termen SNPs som är en förkortning av engelskans Single Nucleotide Polymorphisms. En SNP är en typ av punktmutation.
- En insertion är också typ av punktmutation som innebär att enstaka baspar lagts till.
- En deletion innebär att enstaka baspar, eller större delar av en kromosomen, har försvunnit. När det rör sig om enstaka nukleotider är det också en punktmutation.
- En inversion innebär att delar av kromosomen har hamnat åt fel håll så att DNA-sekvensen blir bakvänd.
- En translokation innebär att en bit av en kromosom bytt plats och hamnat på en annan kromosom.
- En duplikation innebär att en bit av kromosomen kopierats upp så att den upprepas. Uppstår många kopior av samma sekvens kallas det för kopietalsvariation (på engelska Copy Number Variation, som förkortas CNV).
De största mutationerna innefattar hela kromosomer. Ett välkänt exempel är trisomi på kromosom nummer 21 som ger Down syndrom. Det finns då tre istället för två kopior av kromosomen nummer 21. Om alla kromosomer finns i fler än två kopior, till exempel tre (triploid) eller fyra (tetraploid) överlever oftast inte organismen. Ett undantag är växter där olika former av polyploidi är ganska vanligt utan att individen tar skada. Även en del fiskar och salamandrar är polyploida.
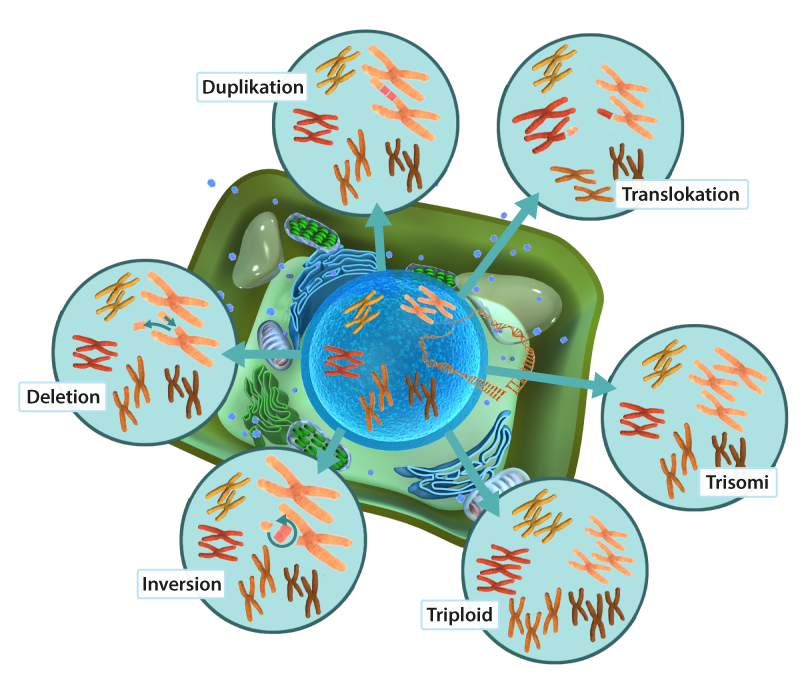
En mutations effekten beror på var i genomet den uppstår
De delar av en gen som kodar för ett protein heter exoner. Om en mutation uppstår i en exon kan den leda till att proteinet byggs upp annorlunda. En ny aminosyra kan då monteras in i proteinet som ger det annorlunda egenskaper, som kan vara till det bättre eller sämre för individen. En mutation i en exon kan också introducera en för tidig stopp-signal som gör att proteinet blir ofullständigt, eller inte tillverkas alls.
De delar av en gen som inte kodar för ett protein heter introner. När en gen läses av (transkriberas) och en arbetskopia i form av ett mRNA bildas finns i det första steget intronerna med. Innan mRNA-molekylen transporteras till ribosomerna för att fungera som mall vid proteintillverkningen, klipps alla introner bort. Den processen kallas för splitsning, eller splicing på engelska. En ”splice site” är den plats i genen som motsvarar ”klipp-ställena” i mRNA-molekylen. En mutation vid ett sådant ställe (en splice site mutation) kan leda till en ofullständig splitsning och att en eller flera introner felaktigt ingår i den färdiga mRNA-molekylen. Proteinets uppbyggnad blir då förändrat.
Mutationer i introner eller i regionerna mellan gener, får vanligtvis ingen effekt alls men det finns undantag. Om en mutation uppkommer på en plats i genomet som styr genuttryck, till exempel i en promotor, kan den påverka hur mycket protein som tillverkas.
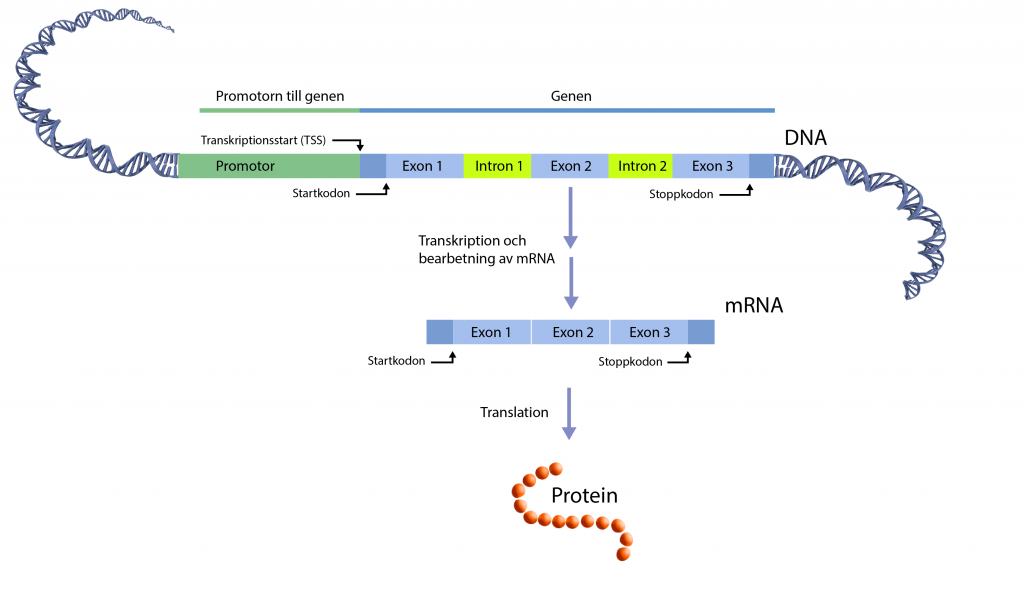
Mutationer påverkar inte alltid proteinets uppbyggnad
I styckena ovan delas mutationer in beroende på hur de ser ut och var de uppstår i genomet. Ett annat sätt att dela in dem är genom att benämna hur de påverkar själva aminosyrasekvensen och därmed proteinets uppbyggnad. Tre och tre bildar nukleotiderna i DNA, som vid proteintillverkningen representeras av mRNA, ett så kallat kodon. Varje kodon översätts med en aminosyra.
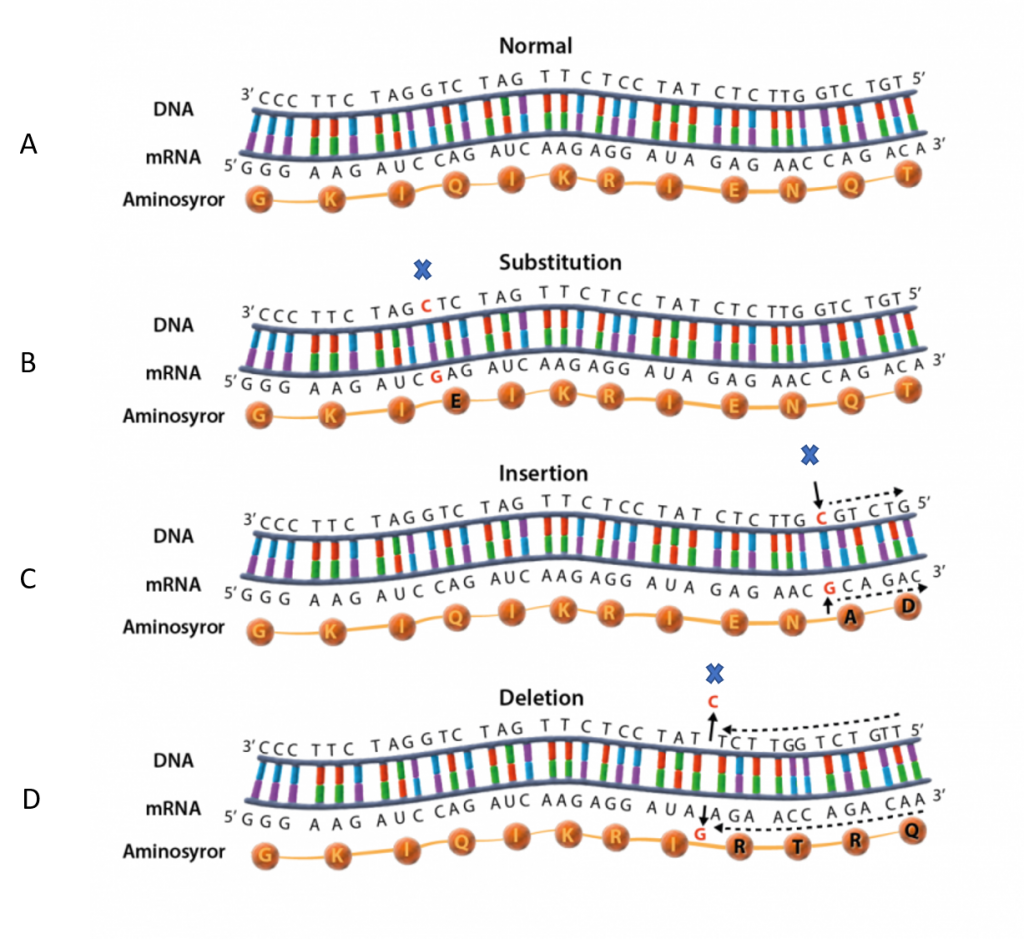
En tyst mutation märks inte
En tyst mutation ändrar DNA- men inte aminosyrasekvensen. Det kan ske eftersom flera kodon översätts med samma aminosyra. Till exempel översätts både kodon AGU och AGC med aminosyran serin. En punktmutation som ändrar U till C påverkar därför inte aminosyrasekvensen och proteinet förblir oförändrat. Mutationen märks alltså inte.
Missense och nonsense ändrar aminosyrasekvensen
Om en punktmutation istället leder till att aminosyrasekvensen förändras kallas den för en missense-mutation. En sådan mutation ändrar proteinets uppbyggnad och kanske dess funktion beroende på vilken del av proteinet som förändrats. Punktmutationer kan även vara nonsense-mutationer som ändrar ett kodon för en aminosyra till ett stopp-kodon. Detta leder till att tillverkningen avslutas för tidigt och proteinet blir förkortat.
En frameshift mutation ändrar läsramen
Om en mutation innebär att nukleotider läggs till eller tas bort i en exon, kan det också innebära att aminosyror läggs till eller tas bort från proteinet. Det sker när antalet nukleotider som omfattas av mutationen är delbart med tre och motsvarar ”jämna” kodon. Om antalet inte är delbart med tre så sker istället en förskjutning av hela läsramen och alla kodon som följer blir förändrade. En sådan mutation kallas för en frameshift indel som på svenska ungefär betyder läsramsförskjutning.

Vid sexuell förökning gynnas den genetiska variationen på flera sätt. Dels genom det uppenbara i att föräldrarnas DNA blandas, men även eftersom nya kombinationer av alleler uppkommer när könsceller bildas. Den celldelningsprocess som ger nya könsceller heter meios men kallas också för reduktionsdelning, eftersom antalet kromosomer halveras. Ur en somatisk (diploid) cell bildas fyra (haploida) könsceller. Eftersom varje könscell är haploid kan två könsceller som smälter samman vid befruktningen bilda en ny diploid cell som är början på ett nytt liv.
Innan den diploida cellen börjar dela sig under meiosen kopieras alla kromosomer. När det är klart ligger kromosomerna uppradade bredvid varandra innan de dras isär och separeras till olika könsceller. I det här steget uppstår nya kombinationer av alleler på två sätt: oberoende nedärvning och homolog rekombination, som leder till att varje könscell blir unik. Det är förklaringen till att helsyskon kan vara ganska olika varandra.
Homolog rekombination (överkorsning)
När kromosomerna kopierats under meiosen och de ligger uppradade inför celldelningen, sker ett utbyte av DNA mellan homologa kromosompar. Det kallas för homolog rekombination eller överkorsning, och leder till en omflyttning av alleler. Under överkorsningen kan utbytet av DNA vara rent fysiskt och innebära att DNA-sekvenser mellan homologa kromosomer klipps ut och byter plats. En DNA-sekvens kan också kopieras från den ena kromosomen och klistras in i den andra, utan att den kopierade strängen ändras.
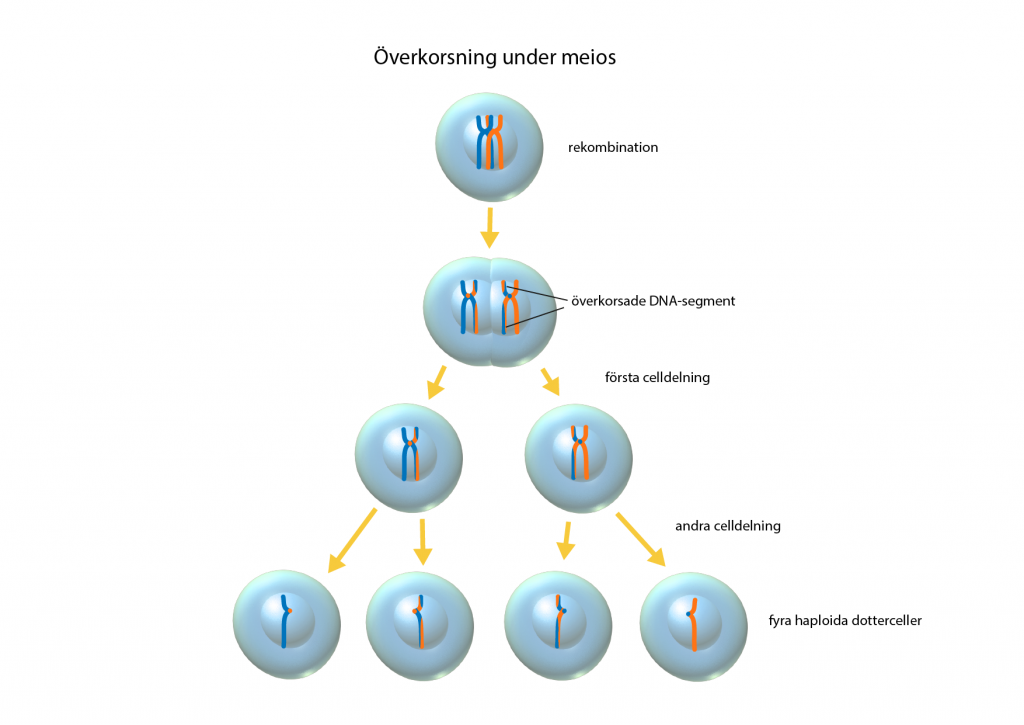
Oberoende nedärvning
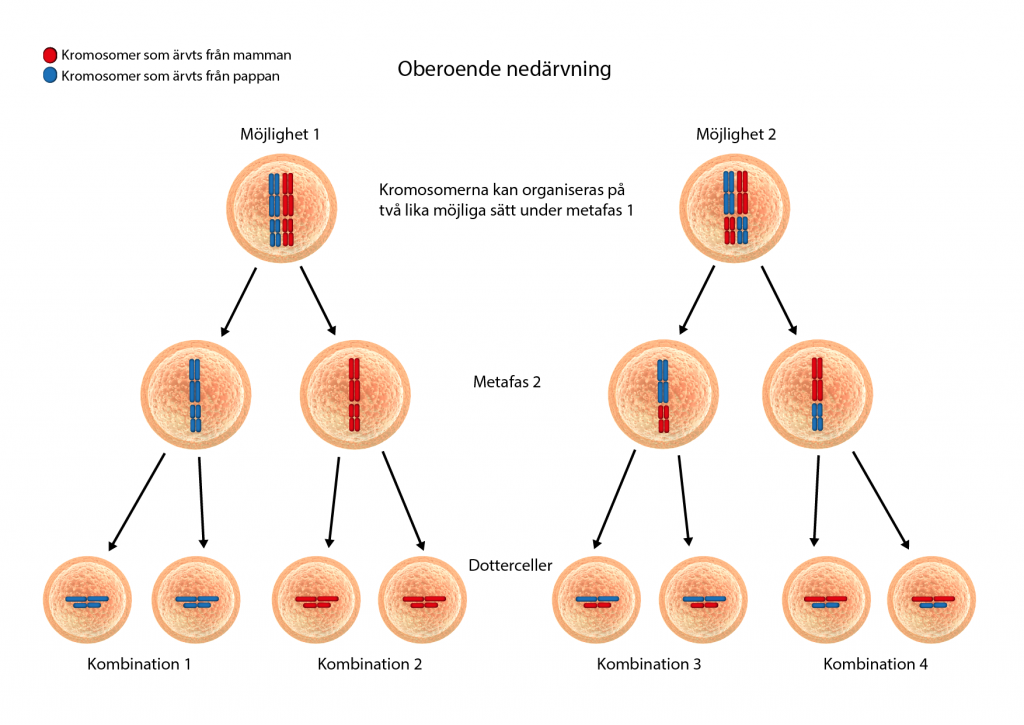
Efter överkorsningen dras kromosomerna isär och separeras till var sin könscell. Vilken kromosom, den som ärvts från mamman eller pappan, som hamnar i vilken könscell är helt slumpmässigt. Det kallas för oberoende nedärvning. Det innebär att två egenskaper som kodas av gener som finns på två olika kromosomer nedärvs helt oberoende av varandra. Det kan märkas i avkomman genom att helt nya kombinationer av egenskaper som inte fanns hos föräldrarna kan uppkomma.
Plats-specifik rekombination (som inte sker under meios)
Omflyttningar av alleler kan också ske i specifika delar av genomet och gynna genetisk variation. Det kallas för plats-specifik rekombination.
V(D)J rekombination trimmar immunförsvaret
Ett exempel är på plats-specifik rekombination är V(D)J-rekombination. Det sker kontinuerligt när vissa immunceller bildas för att göra immunförsvaret snabbare och mer effektivt i att bekämpa till exempel virus. V(D)J-rekombination förekommer bara hos ryggradsdjur och är ett signum för den adaptiva delen av vårt immunförsvar – det som minns infektioner vi redan haft.
Till det adaptiva immunförsvaret hör bland annat B- celler och T-celler. B-celler bildar antikroppar som känner igen främmande ämnen, så kallade antigen, i kroppen. Ett antigen kan komma från till exempel virus, bakterier eller från egna celler som inte beter sig normalt, till exempel cancerceller. T-celler har receptorer på ytan som också känner igen antigen. När en igenkänning sker så aktiveras en immunsvar som attackerar infektionen eller cancercellerna.
När B-celler och T-celler bildas sker en omkastning av de gensegment som kodar för T-cellsreceptorernas och antikropparnas igenkänningsytor. Segmenten heter V (variable), D (diversity) och J (joining), därav benämningen V(D)J rekombination. På det här sättet kan T-celler och antikroppar upptäcka många olika antigen och immunförsvaret blir snabbare och effektivare.
Transposition
Transposition är en annan typ av rekombination som ökar den genetiska variationen. Egentligen är transposition en typ av mutation som innebär att rörliga DNA-sekvenser, så kallade transposoner, fogar in sig på nya platser i genomet. Om en transposon infogas i en gen ger det ofta negativa konsekvenser. Ett undantag är en handfull gener som verkar ha sitt ursprung i transposoner, till exempel generna Rag1 och Rag2 som kodar för de enzymer som driver V(D)J rekombination (som beskrivs i stycket ovan).
Den genetiska variationen i en population ökar också när nya individer flyttar in och för med sig nya alleler. Det kallas genflöde eller migration. Det kan vara en fågel som häckar på ett nytt ställe med nya artfränder, eller ett pollenkorn som färdas med vinden från en ö till en annan. När en individ från population A för med sig nya alleler till population B ökar den genetiska variationen inom populationen B. Samtidigt minskar den genetiska variationen mellan population A och B som blir mer lika.
Faktorer som minskar genetisk variation
Genetisk drift
Större populationer har oftast mer genetisk variation än små. Det beror bland annat på att små populationer har en ökad risk att slumpmässigt förlora genetisk variation över tid. Det kallas för genetisk drift och betyder förenklat att ovanliga alleler försvinner över tid, och vanliga fixeras. När alleler fixeras betyder det att alla individer i en population har samma allel och ingen variation alls förekommer vid detta locus.
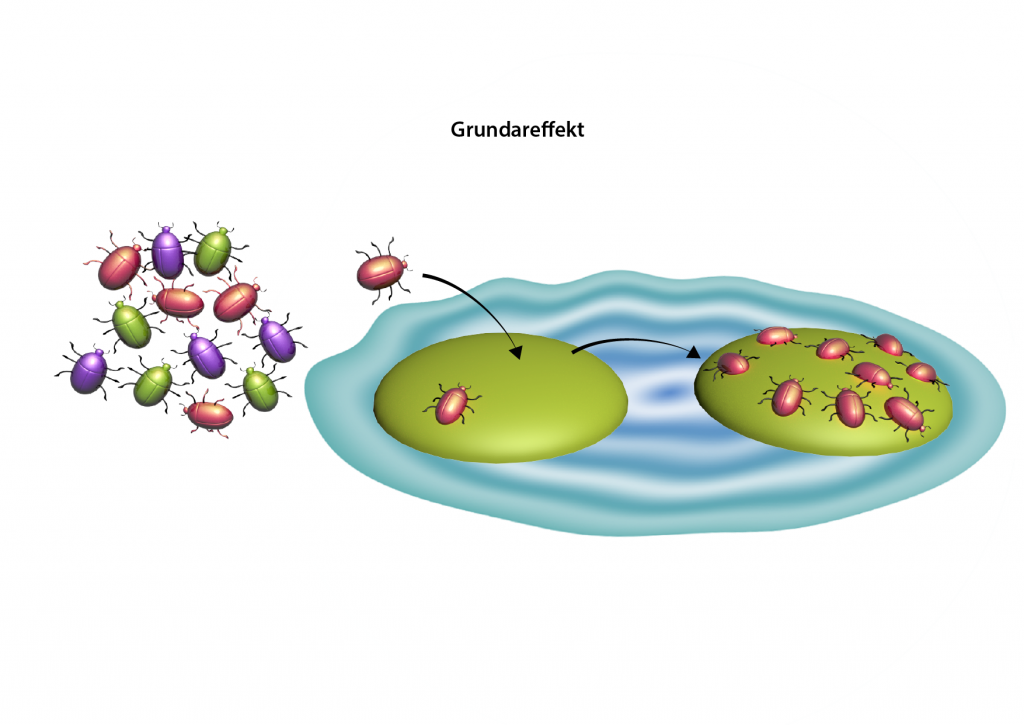
Flaskhals- och grundareffekt
Om antalet individer i en population minskat drastiskt till följd av till exempel en miljökatastrof eller ett sjukdomsutbrott, så har populationen även förlorat genetisk variation. Det kallas för en flaskhalseffekt. Om antalet individer understiger ’minsta livskraftiga population’ så dör den på sikt ut. Om den istället återhämtar sig består den genetiska variationen i populationen av de alleler som fanns hos överlevarna. På lång sikt kan populationen få högre genetisk variation igen med mutationer och genflöde från andra populationer.
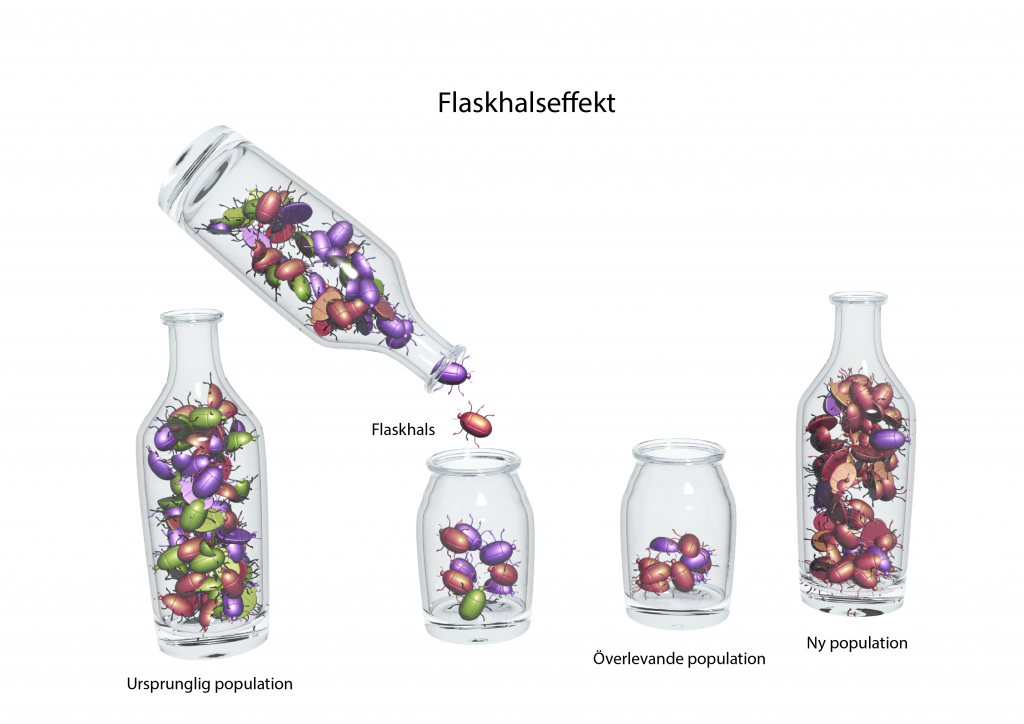
På ett liknande sätt har en ny population som grundats av ett fåtal individer också låg genetisk variation. Det kallas just för grundareffekt och sker exempelvis om ett par fåglar, insekter, eller ett frö från en växt, koloniserar en ny ö där arten inte tidigare fanns.
Små populationer och inavel
I en liten population är också inavel vanligare. Inavel innebär att genetiskt lika individer får en gemensam avkomma. Dels kan inavel leda till en fixering av vanligare alleler och därmed minskad genetisk variation, men också att skadliga recessiva alleler blir vanligare och påverkar fenotypen. Dessa kan orsaka genetiska sjukdomar eller defekter och skulle lättare ha försvunnit ur en stor population där trycket från det naturliga urvalet är starkare.